From the Editor: An Outsider's Perceptions
The Gaze of Animal Life
In the News
Conducting Dog Feeding Trials on the Antarctic Huskies:
a behind the scenes look at how it got done!
Further Experiments on the nutrition of sledge dogs
How Use of the name Inuit became official
An Examination of Traditional Knowledge:
the case of the Inuit Sled Dog, part 4
Chinook Project visits Northern Labrador
Media Review: Qimmit - A Clash of Two Truths
IMHO: In Transition
Index: Volume 12, The Fan Hitch
Navigating This Site
Index of articles by subject
Index of back issues by volume number
Search The Fan Hitch
Articles to download and print
Ordering Ken MacRury's Thesis
Our comprehensive list of resources
Talk to The Fan Hitch
The Fan Hitch home page
ISDI home page
Editor-in-Chief: Sue Hamilton
Webmaster: Mark Hamilton
Print Edition: Imaged and distributed by the IPL students of the Ulluriaq School, Kangiqsualujjuaq, Nunavik
The Fan Hitch,
Journal of the Inuit Sled Dog International,
is published four times a year. It is
available at no cost online at:
https://thefanhitch.org.
Print subscriptions: in Canada $20.00, in USA $23.00, elsewhere $32.00 per year, postage included. All prices are in Canadian dollars. Make checks payable in Canadian dollars only to "Mark Brazeau", and send to Mark Brazeau, Box 151 Kangiqsualujjuaq QC J0M 1N0 Canada. (Back issues are also available. Contact Sue Hamilton.)
The Fan Hitch welcomes your letters, stories, comments and suggestions. The editorial staff reserves the right to edit submissions used for publication.
Contents of The Fan Hitch are protected by international copyright laws. No photo, drawing or text may be reproduced in any form without written consent. Webmasters please note: written consent is necessary before linking this site to yours! Please forward requests to Sue Hamilton, 55 Town Line Rd., Harwinton, Connecticut 06791, USA or mail@thefanhitch.org
Print subscriptions: in Canada $20.00, in USA $23.00, elsewhere $32.00 per year, postage included. All prices are in Canadian dollars. Make checks payable in Canadian dollars only to "Mark Brazeau", and send to Mark Brazeau, Box 151 Kangiqsualujjuaq QC J0M 1N0 Canada. (Back issues are also available. Contact Sue Hamilton.)
The Fan Hitch welcomes your letters, stories, comments and suggestions. The editorial staff reserves the right to edit submissions used for publication.
Contents of The Fan Hitch are protected by international copyright laws. No photo, drawing or text may be reproduced in any form without written consent. Webmasters please note: written consent is necessary before linking this site to yours! Please forward requests to Sue Hamilton, 55 Town Line Rd., Harwinton, Connecticut 06791, USA or mail@thefanhitch.org
The Inuit Sled Dog International
The Inuit Sled Dog International (ISDI) is a consortium of enthusiasts whose goal is the preservation of this ancient arctic breed in its purest form as a working dog. The ISDI's efforts are concentrated on restoring the pure Inuit Dog to its native habitat. The ISDI's coordinators welcome to your comments and questions.
ISDI Coordinator Canada:
Geneviève Montcombroux, Box 206, Inwood, MB R0C 1P0; gmontcombroux@gmail.com
ISDI Coordinator USA:
Sue Hamilton, 55 Town Line Road, Harwinton, CT 06791, mail@thefanhitch.org
Further Experiments on the nutrition of sledge dogs
By H. T. Wyatt
Division of Human Physiology, National Institute for Medical Research,
Medical Research Council, Hampstead, London, N.W. 3
(Received 9 August 1961 – Revised 25 April 1963)
Sledge dogs are still widely used as draught animals on polar expeditions despite the increasing use of mechanical transport. They live and thrive whilst kept continuously out of doors in all but the most extreme conditions. At the bases they are fed on a diet of seal or other raw meat, but during sledging journeys, when fresh meat may not be available, all dog food must be carried on the sledges or previously laid ahead of the parties in depots. For many years the standard British ration was 454 g (1 lb)/day of pemmican, a dehydrated concentrated food developed by Watkins (1932) for the British Arctic Air Route Expedition, but this diet has been considered unsatisfactory both in quantity and quality (Bingham 1941).
Experiments on the nutrition of sledge dogs were conducted both in Britain and in Antarctica (Taylor, Worden & Waterhouse, 1959) and an assessment was made of dog pemmican, which consisted of approximately two-thirds protein and one-third fat. The ration of 1 lb/day was not sufficient to maintain the weight of tethered dogs, and a thiamine deficiency was demonstrated. Diarrhoea was usual, and faecal nitrogen excretion was 30% of the amount ingested. Taylor et al. (1959) developed an new diet for dogs containing approximately 30% protein, 40% fat and 10% carbohydrate, to which were added 1.2 mg thiamine, 10 mg nicotinic acid, 3 mg riboflavin, 1000 i.u. vitamin A, 600 i.u. vitamin D, 20 mg calcium pantothenate and 0.4 mg pyroxidine for each 1 lb block.
Feeding trials were conduced in Antarctica in 1957 and 1958, when the old diet (diet A) was compared with the new diet (diet B) and with a further slight modification of the new diet (diet C). The results of the experiments are reported in this paper.
METHODS
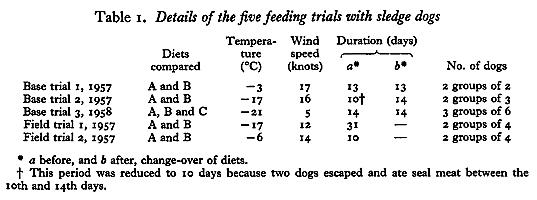
Thirty-two male husky dogs were used at bases (lat. 66º 52’ S, long. 66º 48’ W) and E (lat. 68º 11’ S, long. 67º 00’ W) of the British Antarctic Survey, formerly the Falkland Islands Dependencies Survey. Sixteen were less than 2 years old, twelve between 2 and 6 years old, and four older than 6 years; most had sledged considerable distances before the tests and had been accustomed to concentrated diets while traveling. Before each trial they were fed on seal meat for at least 2 weeks.
Diets A and B were compared in two trials with dogs tethered on snow near the base hut, and in two trials with dogs pulling a loaded sledge in the daytime and tethered separately at night. A third trial with tethered dogs was performed at the base to compare diets A, B and C.
Groups of dogs were selected and matched by weight and if possible by age. Each group was given daily 1 lb/dog of a different diet for periods of up to 31 days and body-weight changes and faecal excretions for N [nitrogen] and fat were measured. In the experiments on tethered dogs the diet given to each group was changed at the mid-point, so that a cross-over experiment was performed; in the trials using working dogs, there was no cross-over. Details of the five trials are given in Table 1:
A spring balance graduated in lb and reading 200 lb (91 kg) was used for measurement of body-weight. Oscillations of the scale pointer were sometimes violent when the balance was suspended from tripod out of doors. Errors were assessed by making ten weighings of one dog within a ½ h period; it was concluded that the weight could be measured to within +/- 1 lb.
On four to six occasions during each trial faeces were collected for 24 h. Errors arising from over- or under-collection of representative 24 h samples, due to irregular defaecation by the dogs and to their almost invariable coprophagy, were minimized by regular hourly inspection of the dog spans. Samples of about 50 g were dried over a boiling water-bath for 10 h, with small additions of absolute alcohol to assist drying. The dried samples were sealed in corked glass tubes and sent to Cambridge where they were further dried to constant weight and analysed for N by the micro-Kjeldahl method with copper selenite as catalyst, and for fat by the method described by King (1946).
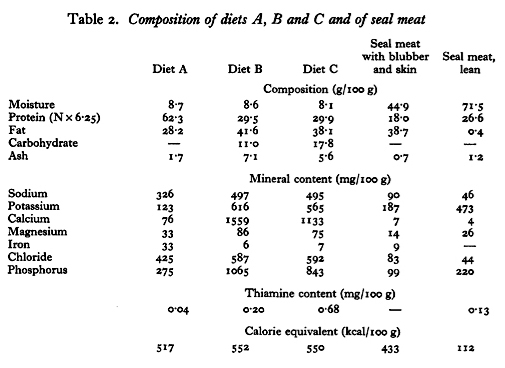
The compositions of the three experimental diets and specimens of lean seal meat, and seal meat including blubber and skin, are shown in Table 2. The methods used for the analysis of these foods were in general those described by McCance and & Shipp (1933) and McCance, Widdowson & Shackleon (1936). The starch in diets B and C was hydrolysed with Takadiasase (Parke, Davis % Co.) and the glucose determined by the Munsen-Walker method (Association of Official Agricultural Chemists. 1950). Sodium and potassium were determined in an EEL flame photometer (Evans Electroselenium Ltd) after phosphate had been removed from the extract of the ash with solid Ca(OH)2 (Widdowson & Southgate, 1959). Thiamine was measured by the thiochrome method of Harris & Wang (1941).
RESULTS
Weight changes
Weight changes
All weight changes were adjusted to a period of 10 days and the means for each trial are shown in Table 3. Weight loss appeared to be greater in the first part of each trial, before the diets given to each group were changed over, than in the second part. No effect due to the change of diet was demonstrated.
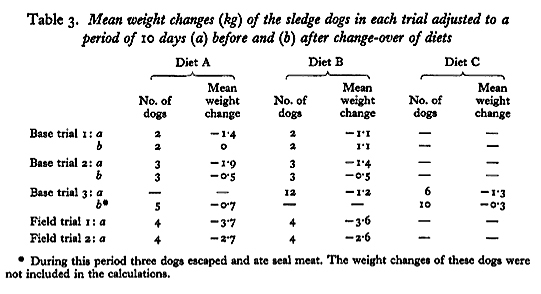
Composition of faeces
The faeces of dogs fed on diet A were noticeably different from those of dogs fed on diets B and C. On diet A the dogs tended to defaecate three to four times daily, producing large amounts of loose, watery, dark faeces. On diets B and C they usually defaecated once or twice daily producing well-formed, light-coloured faeces.
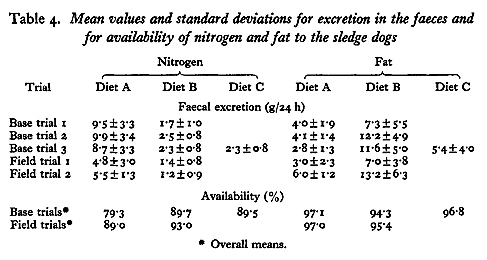
The amounts of N and fat excreted in the faeces, shown in Table 4, differed between diets. Dogs fed on diet A excreted more N and less fat than did dogs fed on diet B.
Utilization of food constituents
To compare the utilization of the food constituents in the three diets the availabilities, i.e. the net absorption as a percentage of the intake, of N and of fat were calculated. The overall means of availability in tethered and in working dogs are shown in Table 4. Comparisons between diets were tested on a within-dog basis.
In tethered dogs the availability of N was greater on diets B and C than on diet A (P <0.001), and in working dogs it was greater on diet B than on diet A (P <0.05). When the dogs were tethered the availability of the fat in diets A and C was greater than in diet B (P < 0.001).
Differences in the availabilities of the food constituents to tethered and to working dogs were tested on a between-dog basis, and were approximate, as no account was taken of the weight-matching of the groups. In working dogs N availability was greater than in tethered dogs for both diet A (P <0.001) and for diet B (P <0.01). There was no evidence of any difference in the availability of fat to tethered and to working dogs fed on either diet.
Performance
Toward the end of the first field trial one dog, Bodger, who had been fed on diet A, refused his food, became listless and apathetic, and began to vomit continuously. When removed from the trial and fed on seal meat he recovered within 48 h. Such episodes did not occur with dogs eating diets B and C.
DISCUSSION
Diets B and C were better utilized than diet A, but 1 lb/day was inadequate to maintain the weight of tethered dogs, and working dogs lost weight rapidly when fed on any of the three diets (Fig. 1). Diarrhoea, which was constant in dogs fed on diet A, did not occur in dogs fed on diet B or C:
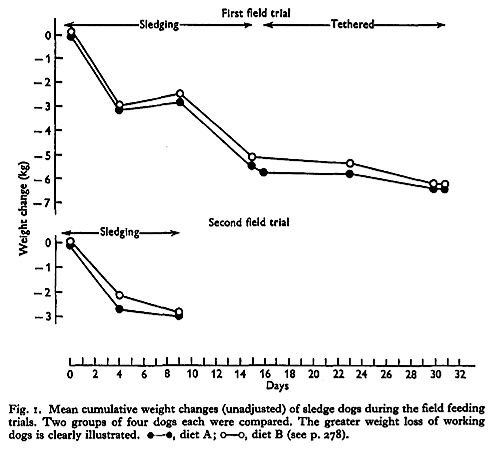
No differences were found in the weight changes of dogs given the old or the new diets. Since the difference in available calories between diet A and diet B or diet C was about 300 kcal/day, differences in weight loss of the order of 40 g/day might have been expected, but such differences would undoubtedly be obscured by the insensitivity of the weighing technique. Differences in water balance on the three diets may also have masked differences in weight change. Taylor et al. (1959) deduced from experiments in Britain that water retention occurred in dogs given diet A, and water requirements on this diet were possibly large so as to replace water loss in the faeces. On diets B and C faecal loss of water was greatly reduced, but because the protein source was fish meal these diets contained large amounts of inorganic salts. It is therefore likely that water requirements were large on all the diets.
The large loss of N in the faeces of dogs fed on diet A, reported by Taylor et al. (1959), was confirmed in these experiments, and the utilization of N by dogs fed on diets B and C was shown to be much better. The high protein intake of dogs fed on 1 lb/day on diet A is not likely to have caused the large faecal N loss, because dogs fed on seal meat take almost the same amount of protein and rather more fat, but excrete in the faeces only about 7% of ingested N (Orr, 1963). The dogs receive 2-3 kg of seal meat and blubber on alternate days; they usually eat all of the meat and some of the blubber, taking more blubber in the winter months and leaving large amounts in the summer. The proportions of meat and blubber vary according to the way in which the meat is cut, and the amounts used in the analyses for Table 2 were arbitrary. Nevertheless, on seal-meat diet dogs must take up to 250 g protein each day, a variable amount of fat, and about 2 mg thiamine.
Various heat treatments of protein have been shown to reduce its digestibility (Mendel & Fine, 1912; Morgan & Kern, 1934; Mader, Schroeder & Smith, 1949; Bissett & Tarr, 1954; Harms & Scott, 1956; Schroeder, Iacobellis & Smith, 1961). Since the protein source of diet A was minced beef that had been steam washed, the processing of the meat meal used may have cause the poor protein utilization.
With all the diets the availability of fat was within the normal range, although it was less than with diet B than with either diet A or C. In husky dogs, Taylor et al. (1959) found that the faecal fat amounted to 3-7% of the ingested fat, where as for men eating a high-fat diet polar regions Butson (1950) reported that faecal fat was 7% of the dietary intake, and Masterton, Lewis & Widdowson (1957) found that it was 3-4% of the ingested fat during sledging journeys.
Since the availability of N was greater for working than for tethered dogs, whereas the availability of fat remained with in the normal range, it appears that the diet was better utilized during hard work.
Diet B has been used by the Canadian Defence Research Board, by the Trans-Antarctic Expedition and by the British Antarctic Survey. Although comparison of the performance of dog teams fed on different diets is complicated by the fact morale and interest or boredom play a large part (Taylor, 1957a), dog drivers on these expeditions have reported that dogs perform better on diet B than on diet A.
The figures obtained in the feeding trials can be used to estimate the energy requirements of sledge dogs. The daily calorie equivalents of the foodstuffs available to tethered dogs were (from Table 3): from diet A 2080 kcal, from diet B 2360 kcal, and from diet C 2340 kcal. Since the dogs still lost weight it is evident that they required over 2360 kcal/day to maintain body-weight. It has been calculated that the maximum useful external work which a sledge dog can perform in 1 h is 120 kcal and in a normal day’s travel between 250 and 400 kcal (Taylor, 1957b). These figures represent a calorie requirement of 1000-1600 kcal/day if the efficiency of muscular work is taken to be as high as 25% (Hill, 1939, 1950). If it be assumed that the calorie requirements for external work were additional to the requirements of tethered dogs, the total calorie requirement of a working dog probably lies between 3500 and 4600 kcal/day. Large as this requirement seems for an animal between 35 and 45 kg body-weight, the weight losses of working dogs seem to support the estimate (Fig 1.).
It is thus improbable that a 1 lb block of concentrated food can be provided that would be nutritionally adequate in all respects, since 1 lb of fat daily would provide barely sufficient energy for working sledge dogs.
SUMMARY
1. Thirty-two husky dogs were fed on three different concentrated diets in the Antarctic. Feeding trials, during which each dog was given 1 lb/day of one or other diet, were performed both during sledging journeys and while the dogs were kept tethered and unworked.
2. When fed on a diet containing 30% protein, 40% fat and 10-20% carbohydrate the dogs excreted much less nitrogen and tended to loose less weight than on a diet containing approximately 66% protein, and 33% fat. The large amounts of N excreted by dogs fed on the diet containing 66% protein may be related to the source of protein ingredients rather than to the absolute amounts of protein. Fat availability on both diets was within the normal range.
3. The evidence suggests that the energy expenditure of working sledge dogs, weighing 35-45 kg, is of the order of 4000 kcal/day, and may be greater.
I thank Dr E. M. Widdowson for criticism and advice and for arranging for the analyses of the foodstuffs and the faeces, Mrs M. P. Williams for performing the analyses, Dr E. Kodicek for the analyses of thiamine, Miss P. Howlett for statistical advice and Lt-Cdr A. B. Erskine, R.N., and Mr K. Hoskins for invaluable assistance and encouragement under trying conditions in the Antarctic.
REFERENCES
Association of Official Agricultural Chemists (1950). Official Methods of Analysis, 7th ed. Washington, D.C.: Association of Official Agricultural Chemists.
Bingham, E. W. (1941). Polar Rec. 3, 367.
Bissett, H. M. & Tarr, H. L. A. (1954). Poult. Sci. 33, 250.
Butson, A. R. C. (1950). Lancet, 258, 993.
Harms, A. J. and Scott, P. P. (1956). J. Sci. Fd Agric. 7, 477.
Harris, L. J. & Wang, V. L. (1941). Biochem. J. 35, 1050.
Hill, A. V. (1939). Proc. Roy. Soc. B, 127, 434.
Hill, A. V. (1950). Sci. Progr. Twent. Cent. 38, 209.
King, E. J. (1946). Micro-analysis in Medical Biochemistry. London: J. and A. Churchill.
McCance, R. A. & Shipp, H. L. (1933). Spec. Rep. Ser. med. Res. Coun., Lond., no. 187.
McCance, R. A., Widdowson, E. M. & Shackleton, L. R. B. (1936). Spec. Rep. Ser. med. Res. Coun., Lond., no. 213.
Mader, I. G., Schroeder, L. J. & Smith, A. H. (1949). J. Nutr. 39, 341.
Masterton, J. P., Lewis, H. E. & Widdowson, E. M. (1957). Brit. J. Nutr. 11, 346.
Mendel, L. B. & Fine, M. S. (1912). J. biol. Chem. 11, 5.
Morgan, A. F. & Kern, G. E. (1934). J. Nutr. 7, 367.
Orr, N. W. M. (1963). J. Physiol. 165, 51 P.
Schroeder, L. J., Iacobellis, M. & Smith, A. H. (1961). J. Nutr. 73, 143.
Taylor, R. J. F. (1957a). Polar Rec. 8, 317.
Taylor, R. J. F. (1957b). J. Physiol. 137, 210.
Taylor, R. J. F., Worden, A. N. & Waterhouse, C. E. (1959). Brit. J. Nutr. 13, 1.
Watkins, H. G. (1932). Geogr. J. 79, 466.
Widdowsen, E. M. & Southgate, D. A. T. (1959). Biochem. J. 72, 200.